CITOLOGÍA
...PARA COMENZAR...
A LEER Y TOMAR APUNTES !!!! :)
CITOLOGÍA

Las células se han clasificado, de acuerdo a la presencia o ausencia de núcleo verdadero, en dos grandes grupos: células procariontes y células eucariontes.
Células procariontes: son aquellas en las que el núcleo se encuentra difuso en el citoplasma, es decir, son las que no poseen un núcleo celular rodeado por una membrana (pro = antes de, karyon = núcleo).
Células eucariontes: son aquellas que poseen un núcleo celular delimitado por una doble membrana (eu = verdadero, karyon = núcleo).
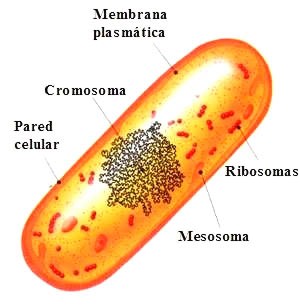 |
Célula procarionta o procariota
|
Células procariontes o procariotas son la forma de vida más simpleque se conoce, en cuanto a estructura y función.
Célula cuyo material genético ni se organiza en un núcleo bien definido ni se reparte durante la reproducción celular.
Los seres procariontes son siempre unicelulares y pertenecen al reino de los moneras, como las bacterias y las algas verde-azuladas.
El método de replicación de los procariontes se denomina fisión binaria, y consiste en una replicación del ADN (expresado en un solo cromosoma) seguida de una duplicación celular. Como no poseen núcleo diferenciado no pueden utilizar mitosis como su forma de reproducción, que es exclusiva para los eucariontes.
Existen organismos procariontes de muy variadas formas: hay organismos esféricos, con forma de bastón, con forma espiralada y con forma ovoide.
Aunque su forma externa sea diferente su composición interna es muy parecida. En general poseen una cubierta externa protectora llamadapared celular, bajo la cual se encuentra la membrana plasmática, que tiene por función intercambiar sustancias entre la célula y el medio que la rodea y delimitar, además, al citoplasma o citosol donde ocurren todos los procesos químicos que permiten el desarrollo y funcionamiento de la célula.
En las células procariontes, el material genético (DNA) se encuentra libre en el citoplasma sin ninguna estructura que lo delimite, es decir, se halla difuso.
 |
Otra característica que presentan estas células es que no poseen organelos membranosos.
Las enzimas que permiten la degradación (transformación en sustancias más simples) de lípidos e hidratos de carbono para obtener energía se encuentran en el citoplasma al igual que el DNA y otras estructuras que permiten el funcionamiento de la célula.
Los biólogos postulan que las células procariontes son la línea evolutiva más antigua que se conoce y que de ellas se habrían derivado las células eucariontes.
Ellos creen que una célula procarionte fue capaz de formar un núcleo verdadero y que, posteriormente, esta primitiva célula eucarionte incorporó en su citoplasma a otra célula procarionte de menor tamaño.
Célula eucarionte (eucariota o eucariótica)
Las células eucariotas son generalmente mayores y con una estructura más compleja que las células procariotas. La morfología de estos organismos puede incluir apéndices, pared celular, membrana y varias estructuras internas.
Están presentes en células que forman parte de los tejidos de organismos pluricelulares, que pertenecen a los reinos fungi, metazoo y metafíta.
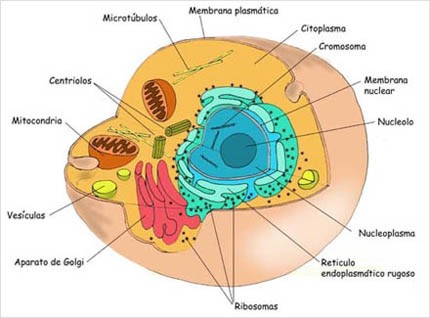 |
Célula eucariota
|
Se caracterizan por tener un núcleo delimitado por una doble membrana (membrana nuclear), que lo separa del resto del citoplasma, donde se almacena el material genético; poseen además organelos membranosos (mitocondrias, lisosomas, cloroplastos, etc).
Poseen formas y tamaños muy variados, de acuerdo a la función que cumpla la célula eucarionte en el organismo.
Las células eucariontes poseen más DNA (ácido desoxirribonucleico) que las células procariontes. El DNA eucarionte se une a proteínas, constituyendo los cromosomas.
Además poseen complejos supramoleculares muy importantes, como es el caso del citoesqueleto, el cual es un verdadero esqueleto interno.
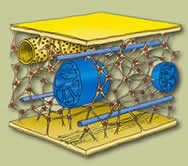 |
Citoesqueleto
|
El citoesqueleto celular consiste en una malla tridimensional de filamentos proteicos cuyas principales funciones son:
• proporcionar el soporte estructural para la membrana plasmática y los orgánulos celulares
• proporcionar el medio para el movimiento intracelular de organelas y otros componentes del citosol
• proporcionar el soporte para las estructuras celulares móviles especializadas, como cilios y flagelos, responsables de la propiedad contráctil de las células en tejidos especializados como el músculo
Hay que tener presente que no todas las células eucariontes presentan los mismos organelos. En las células vegetales y animales es donde se producen las mayores diferencias.
Cilios y flagelos
Algunas células tienen proyecciones del citoesqueleto que sobresalen de la membrana plasmática. Si las proyecciones son pocas y muy largas, reciben el nombre de flagelos.
 |
Espermatozoides
|
El único ejemplo de célula humana dotada de flagelo es el espermatozoide que lo utiliza para desplazarse.
Si las proyecciones son muchas y cortas, se denominan cilios. El ejemplo más típico son las células del tracto respiratorio cuyos cilios tienen la misión de atrapar las partículas del aire.
Al igual que las bacterias, muchas células eucariotas poseen estas estructuras para la locomoción.
Los cilios de las eucariotas son idénticos a los flagelos de las procariotas en estructura, aunque son más cortos y numerosos. Su estructura es más compleja que la de las procariotas, están compuestos por microtúbulos, 9 pares que rodean un par central todo ello rodeado por una membrana.
El flagelo de las eucariotas se mueve como un látigo al contrario de las procariotas que lo hacen rotando como un sacacorchos.
Pared celular
Plantas, algas y hongos poseen pared celular mientras que el resto de las eucariotas no la poseen. La pared celular mantiene la forma celular y previene de la presión osmótica.
La pared celular de las plantas, algas y hongos son distintas a la de las bacterias en cuanto a su composición y estructura física. Por ejemplo, la pared celular de eucariotas no contiene peptidoglucano.
En plantas está compuesta de polisacáridos como la celulosa y pectina. La de los hongos filamentosos contiene quitina y celulosa y en levaduras manano. En las algas existe celulosa, otros polisacáridos y carbonato cálcico.
Membrana citoplásmica o citiplasmática
Independientemente de que la célula eucariota posea o no pared celular, posee membrana citoplasmática que rodea a la parte principal de la célula.
La membrana semipermeable es una bicapa lipídica que posee insertadas proteínas. Algunas de estas proteínas atraviesan enteramente la membrana creando poros a través de los cuales los nutrientes entran dentro de la célula. A estas proteínas se las denomina permeasas.
Las diferencias existentes entre la membrana de eucariotas y procariotas son:
• Los eucariotas contienen esteroles (fundamentalmente colesterol) que le confieren rigidez a la membrana.
• En aquellos eucariotas que no poseen pared celular, la membrana está reforzada por microtúbulos de las proteínas actina y miosina.
• Los eucariotas no localizan los enzimas implicados en la generación de energía metabólica en su membrana.
Las diferencias existentes entre la membrana de eucariotas y procariotas son:
• Los eucariotas contienen esteroles (fundamentalmente colesterol) que le confieren rigidez a la membrana.
• En aquellos eucariotas que no poseen pared celular, la membrana está reforzada por microtúbulos de las proteínas actina y miosina.
• Los eucariotas no localizan los enzimas implicados en la generación de energía metabólica en su membrana.
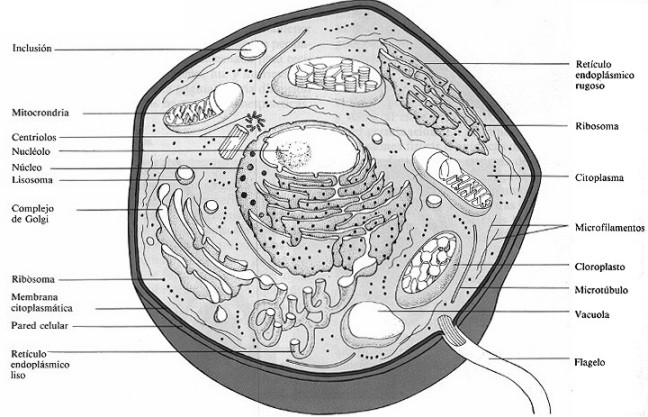
Organelos o argánulos celulares
Dentro de la membrana citoplásmica está el protoplasma que se divide en carioplasma y citoplasma.
El carioplasma es el material que hay dentro de la membrana nuclear, mientras que el citoplasma es el material existente entre la membrana nuclear y la membrana citoplásmica.
En el citoplasma es donde se encuentran los organelos u orgánulos celulares (verdaderas fábricas en miniatura) que son estructuras rodeadas de membrana que realizan funciones especiales, tales como la fotosíntesis y respiración.
Al contrario que las procariotas, el citoplasma de las eucariotas posee una extensa red de microtúbulos y estructuras proteicas que constituyen el citoesqueleto de la célula. Este citoesqueleto genera la forma de la célula y a través de él se mueven los organelos u orgánulos en el citoplasma.
Los organelos u argánulos son:
Núcleo
El núcleo de las eucariotas se caracteriza por su membrana nuclear; es una doble membrana la cual se asemeja a dos membranas citoplasmáticas juntas, que contiene muchos poros grandes a través de los cuales pasan sustancias como proteínas y RNA. Normalmente posee forma esférica u oval.
El núcleo contiene la información hereditaria de la célula en la forma de DNA. En el carioplasma que no se está dividiendo el DNA está combinado con proteínas como las histonas, dándole una apariencia fibrilar. Esta combinación de DNA y proteínas se llamacromatina. Durante la división celular la cromatina se condensa en cromosomas.
Dentro del carioplasma se encuentra el nucléolo, el cual aparece más oscuro con el microscopio electrónico. Alrededor del cinco al diez por ciento del nucléolo es RNA, siendo el resto proteína. Esta estructura es el lugar de síntesis del RNA ribosomal y de los componentes esenciales del ribosoma.
Dentro del carioplasma se encuentra el nucléolo, el cual aparece más oscuro con el microscopio electrónico. Alrededor del cinco al diez por ciento del nucléolo es RNA, siendo el resto proteína. Esta estructura es el lugar de síntesis del RNA ribosomal y de los componentes esenciales del ribosoma.
Los componentes proteicos de los ribosomas sintetizados en el citoplasma entran en el núcleo a través de los poros nucleares para combinarse con el RNA ribosomal recién sintetizado. Tanto las proteínas como el RNA forman las dos subunidades de los ribosomas que salen del carioplasma a través de los poros y se convierten en funcionales en el citoplasma.
Los ribosomas de eucariotas son mayores que los de procariotas.
Retículo endoplásmico
El retículo endoplásmico es una red membranosa de sacos y túbulos que a menudo están conectados a la membrana nuclear y citoplásmica.
 |
En esquema, arriba derecha y abajo izquierda, retículos endoplasmáticos
|
Existen dos formas de retículo endoplásmico: el rugoso y el liso.
El rugoso posee ribosomas y el liso no. Las proteínas sintetizadas en el rugoso son liberadas en el citoplasma o pasan a través de su membrana dentro de los canales por donde son distribuidas a distintas partes de la célula.
El retículo endoplásmico liso está implicado en la síntesis de glucógeno, lípidos y esteroides. Los canales del retículo endoplásmico liso también sirven para la distribución de las sustancias sintetizadas en él.
Aparato de Golgi (o complejo de Golgi)
Está compuesto de sacos membranosos que tienen vesículas esféricas en sus extremos. Fue descrito por primera vez por Camillo Golgi en 1898.
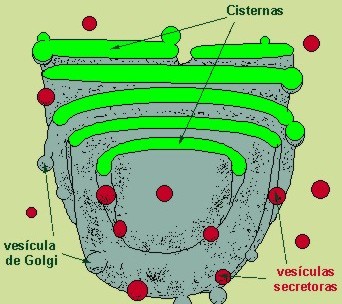 |
Aparato de Golgi
|
Es el centro de empaquetamiento de las células eucariotas, responsable del transporte seguro de los compuestos sintetizados al exterior de la célula.
El aparato de Golgi está conectado a la membrana citoplasmática donde se fusiona y así poder excretar el contenido fuera de la célula, proceso que se llama exocitosis.
Otra función es la de empaquetar ciertos enzimas sintetizados en el retículo endoplásmico rugoso en unos orgánulos llamados lisosomas.
Estos enzimas catalizan reacciones hidrolíticas incluyendo proteasas, nucleasas, glicosidasas, sulfatasas, lipasas y fosfatasas.
El contenido de los lisosomas no se excreta sino que permanece en el citoplasma y participa en la digestión citoplásmica de los materiales ingeridos o absorbidos por la célula.
El que los enzimas hidrolíticos permanezcan dentro del lisosoma protege a la célula de la acción lítica de estos enzimas. En adicción, el aparato de Golgi contiene glicosiltransferasas que unen moléculas de carbohidrato a proteínas para formar glicoproteínas.
Mitocondria
Es un orgánulo citoplásmico donde se generan las moléculas de ATP durante la respiración aeróbica. La membrana interna está muy invaginada y es donde tiene lugar la conversión de energía.
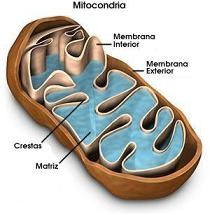 |
Mitocondria
|
Aunque las mitocondrias son orgánulos de células eucariotas se parecen a las células procariotas; contienen sus propios ribosomas, su propio DNA el cual es una única molécula circular que contiene la información genética necesaria para la síntesis de un limitado número de proteínas cuya síntesis tiene lugar en los propios ribosomas de las mitocondrias.
Finalmente, las mitocondrias se dividen para formar nuevas mitocondrias de forma parecida a como lo hacen los procariotas e independientemente del núcleo celular; sin embargo, no se pueden dividir si se sacan del citoplasma.
Cloroplastos
Es el lugar donde ocurren las reacciones fotosintéticas, donde se utiliza la luz como fuente de energía para convertir el CO2 en azúcar y los átomos de O2 del H2O en moléculas de O2 gaseoso.
El cloroplasto es una estructura rodeada por una doble membrana cuyo interior se denomina estroma.
La membrana interna se pliega en el estroma formando sacos en forma de discos llamados tilacoides, los cuales contienen la clorofila y los carotenos que intervienen en la fotosíntesis.
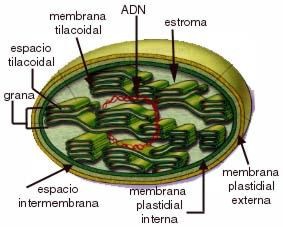 |
Cloroplasto
|
Cada conjunto de tilacoides se llama grano.
Algunos tilacoides se unen a otros de otro grano formando una red.
Los cloroplastos poseen las mismas características que las mitocondrias (ribosomas 70 S, DNA circular, fisión binaria).
La similitud de las mitocondrias y los cloroplastos con los microorganismos procariotas dio base a la teoría endosimbiótica del origen de estos orgánulos.
Bibliografía:
Enciclopedia Interactiva Siglo XXI. Tomo 5
Ciencias Biológicas II. Santillana.
MATERIAL MULTIMEDIAL DE ESTUDIO....
1- VIDEO MUY BÁSICO SOBRE CÉLULAS EUCARIOTAS
ATENCIÓN !!!! TAL VEZ NECESITES INVESTIGAR EN OTROS TEXTOS.....HAY MATERIAL DISPONIBLE EN LA FOTOCOPIADORA DEL COLEGIO.

IREMOS RESOLVIENDO LA SIGUIENTE GUÍA EN FORMA DETALLADA, HAREMOS LA CORRECCIÓN EN LAS CLASES:
1- COMPOSICIÓN, UBICACIÓN Y FUNCIÓN DEL CITOSOL.
2- ¿QUÉ ES EL CITOESQUELETO ?¿CUÁLES SON SUS FUNCIONES?¿QUÉ COMPONENTES DEL CITOESQUELETO COLABORA EN LA FORMACIÓN DE LAS ESTRUCTURAS DE LOS CENTRÍOLOS, LOS CILIOS Y LOS FLAGELOS?
3- COMPOSICIÓN, UBICACIÓN Y FUNCIÓN DEL CENTROSOMA.
4- ¿CUÁL ES LA DIFERENCIA FUNCIONAL ENTRE CILIOS Y FLAGELOS ?
5- COMPOSICIÓN, ESTRUCTURA, UBICACIÓN Y FUNCIÓN DE LOS RIBOSOMAS.
6- ESTRUCTURA, UBICACIÓN Y FUNCIÓN DEL RETÍCULO ENDOPLASMÁTICO LISO Y RUGOSO.
7- ESTRUCTURA, UBICACIÓN Y FUNCIÓN DEL COMPLEJO (APARATO) DE GOLGI.EXPLICA EL PROCESAMIENTO Y ENVOLTURA DE LAS PROTEÍNAS EN ESTE COMPLEJO.
8- ESTRUCTURA, UBICACIÓN Y FUNCIÓN DE LOS LISOSOMAS.
9- ESTRUCTURA, UBICACIÓN Y FUNCIÓN DE LOS PEROXISOMAS.
10- ESTRUCTURA, UBICACIÓN Y FUNCIÓN DE LOS PROTEASOMAS.
11- MITOCONDRIAS. INDICA ESTRUCTURA, UBICACIÓN Y FUNCIÓN .¿QUÉ PROCESOS TIENEN LUGAR EN LAS CRESTAS Y EN LA MATRIZ MITOCONDRIAL?¿POR QUÉ TODOS TENEMOS ADN MITOCONDRIAL EXCLUSIVAMENTE HEREDADO DE NUESTRA MADRE?
12- ESTRUCTURA, UBICACIÓN Y FUNCIÓN DEL NÚCLEO.¿CÓMO HACEN LAS PARTÍCULAS GRANDES PARA ENTRAR Y SALIR DEL NÚCLEO?¿DÓNDE SE PRODUCE EL ARN?¿CÓMO SE DISPONE EL ADN EN EL NÚCLEO?
13- DESCRIBE LA SECUENCIA DE PROCESOS QUE TIENEN LUGAR EN LA SÍNTESIS DE PROTEÍNAS.LUEGO DIFERENCIA TRANSCRIPCIÓN DE TRADUCCIÓN.
14- DIVISIÓN CELULAR. ¿ CUÁLES SON LOS HECHOS MÁS IMPORTANTES DE CADA FASE DE LA MITOSIS? ¿CUÁLES SON LOS HECHOS MÁS IMPORTANTES DE CADA FASE DE LA MEIOSIS?
15- ¿CÓMO SE DIFERENCIAN LAS CÉLULAS?
16- ¿CÓMO ENVEJECEN LAS CÉLULAS?



.preview.jpg)
.preview.jpg)
.preview.jpg)

